![]()
| Introducción a la Teoría de Stephenson |
|||||||||||||||||||||||||||||||||
|
|
|
|
|||||||||||||||||||||||||||||||
|
|
Receptores de reserva |
|
|||||||||||||||||||||||||||||||
|
|
|
|
|||||||||||||||||||||||||||||||
|
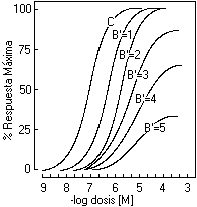
En los ańos 1955, Furchgott, y 1956, Nickerson, en forma simultánea e independiente, demostraron que en algunos tejidos, un bloqueo irreversible de una fracción del "pool" total de receptores presentes, podría producir un desplazamiento paralelo y hacia la derecha de la curva dosis-respuesta de un agonista, que interactuase con estos receptores, antes que se observase una depresión del efecto máximo. Esto sugiere, que al menos en algunos tejidos, es necesario inactivar un elevado porcentaje de los receptores presentes antes que ocurra una disminución de la respuesta máxima. Estos hallazgos se contradicen con los postulados de las teorías anteriores, que según ellas el efecto máximo sólo era obtenido ocupando el 100% de los receptores presentes en el tejido en estudio. Debido a esto, Nickerson propuso en 1956, que en algunos tejidos existe un exceso de receptores, los cuales no necesitan ser activados para generar la respuesta máxima del sistema. Este excedente de receptores se les llamó receptores de reserva. Queda claro, entonces, que la respuesta máxima puede ser producida por algunos agonistas con concentraciones bastante inferiores a aquellas necesarias para saturar la población de receptores. Es necesario enfatizar, que estos receptores de reserva no corresponden a un "pool" diferente, sino que como se mencionó, corresponden a un excedente de los receptores presentes, todos ellos con las mismas características cinéticas y farmacológicas
Este concepto de receptores de reserva implica que pueden existir más receptores que el mínimo necesario para generar la máxima respuesta en un sistema receptor-efector y que por lo tanto no es necesario ocupara el 100% de ellos para inducir el máximo efecto, tal cual había sido postulado por Clark y Ariëns. Es así como en algunos tejidos, es posible inactivar irreversiblemente una elevada fracción de la población total de receptores presentes y aún, bajo estas condiciones, obtener la máxima respuesta. Sólo, cuando el total de receptores viables es inferior al mínimo necesario para inducir la máxima respuesta, ésta se deprime en su magnitud. Por tanto, una máxima ocupación no es el factor limitante que determina la máxima respuesta de un agonista, así como tampoco existe necesariamente una relación lineal entre la fracción de receptores ocupados y el efecto. Como corolario de lo anterior, la DE50 no es una buena aproximación del valor de la Ka, ya que la primera no es necesariamente la concentración de fármaco que ocupa el 50% de los receptores totales, como lo es la Ka. Debido a esto, la constante de afinidad de un agonista por sus receptores no puede ser calculada directamente de su curva dosis-respuesta. De hecho, diferencia de hasta 100 veces se han observado entre ambos valores; por ejemplo, en arteria de conejo Besse & Furchgott en 1976, observaron que el valor de la DE50 fue de 22 nM mientras que el de la Ka de 339 nM, para los efectos contáctiles de la noradrenalina interactuando con a-adrenoceptores. Sus cálculos demostraron que sólo se necesitaba ocupar el 6% de los receptores presentes, en esta preparación, para obtener el 50% del efecto máximo, una concentración de agonista bastante menor a aquella necesaria para ocupar el 50% de los receptores totales (valor de Ka). (siguiente) |
|||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||